Back to my homepage
Back to my homepage |
Chemosensory Orientation in Asterias forbesi
Like many other marine animals, starfish rely heavily on chemoreception in their interactions with their environment. Lacking image-forming eyes and any sense of hearing, chemoreception is their primary sense. The unusual morphology of starfish, however, presents a number of unique problems for orientation, and determining their solutions to these problems was the focus of my work. |
|
 |
Work in the Atema lab (where I did this work) and elsewhere has again and again demonstrated the importance of paired, bilateral sense organs in orientation. Pairs of antenna or antennules as in the lobster, paired rhinophores in the nautilus, and so forth; the paired structures allow the orienting animal to make comparisons of odor distributions in two directions, and this information is often crucial. The pentamerously radial structure of a starfish lacks obvious paired sensors. In fact, it even lacks the simple distinction of a front end: any side of a starfish is equally likely to be the front at any time, at least in the absence of stimulus. |
|
Starfish also lack a central coordinating nervous center — a brain. Instead, a radial nerve runs down the length of each ray, connected with the other radial nerves by a circumoral nerve ring in the body. The nerve ring provides connections, but is not a processing center, and its structure is much simpler than the radial nerves. It appears that one radial nerve can take control of the body, signalling the others to coordinate movement in the direction of its ray. When the starfish moves, this ray is called the leading ray. The leading ray also tends to show a distinct posture, with the tip raised and curled back, and the terminal tube feet actively waving around. In fact, these tube feet look rather like antenna, and probably serve a similar role. The tips of the tube feet are packed with (presumptive) sensory cells. I've conducted two basic types of experiments. In the most straightforward ones, I observed starfish orienting in a large (12 m x 2 m) flume, with and without odor. I've also performed experiments in which I removed the ability to smell from 1, 2, or 3 rays and then observed how orientation behavior changed. (I did this by immersing the tips of the rays in distilled water, which selectively destroys chemoreceptor cells in many marine invertebrates. The starfish recover their sense of smell after a couple days.) The results of both types of experiment suggest that starfish use chemical concentration information (which may be actual concentration or some other correlated parameter, such as the slope or height of odor peaks) to compare concentration between their rays and move in the direction of the highest concentration. Sometimes this moves them upstream, while at other times it moves them across-stream towards the plume axis. As a further test of this theory, I've developed a computer model of a starfish using such a strategy to orient in an odor plume. It is surprisingly reliable at reaching the odor source, and the paths it takes are similar to real orientation paths, both in their routes and in the frequencies with which different rays lead. |
|
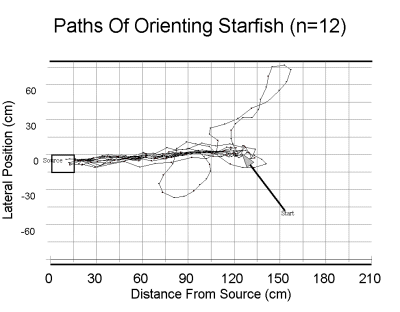
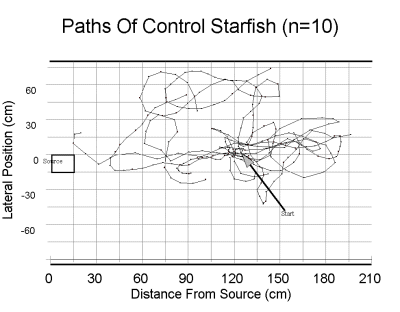
| Above, the paths of twelve starfish in an odor plume. Starfish began (arrow) 120 cm downstream from the source (rectangle at left). As can be seen, the starfish stayed in the plume as they approached the source, with the exception of two starfish which circled before beginning their approach. Below, a similar plot of starfish in the absence of odor. | |
|
|
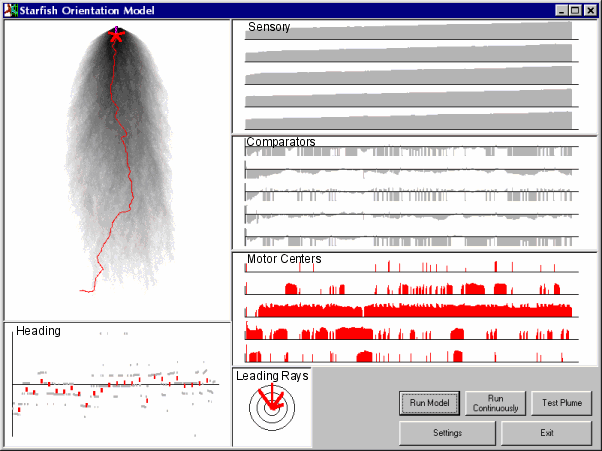 |
| A screen image from the model. The path of the simulated starfish is shown in red against the simulated plume in gray. On the right, plots of the states of the model sensory units, comparators, and motor centers (abstractly representing sensory input, comparisons of sensory input, and motor output). Below, the circular graph shows the frequency with which different rays lead (real starfish mainly led with their upstream or adjacent on the right side rays, or a combination of the two) and on the lower left a plot of mean heading (real starfish showed decreasing headings with approach to the source). Both real and model starfish were variable in their behavior, but model behavior was surprisingly close to that of real starfish. |
Further ReferencesI have two published papers on starfish orientation. "Chemosensory search behavior in the starfish Asterias forbesi" is an account of the paths of orienting starfish. (Dale, J. (1997). Chemosensory search behavior in the starfish Asterias forbesi. Biol. Bull. 193: 210-212.) A second paper, Coordination of chemosensory orientation in the starfish Asterias forbesi, covers leading ray behavior and the lesion experiments. My dissertation is available from UMI or you can contact me directly for a complete copy in PDF format (I'd post it here but it's a bit large with all the figures). If you're looking for more information on starfish behavior, my list of references relevant to starfish behavior research is also available on-line in HTML format.
Contact InformationJonathan Dale can be contacted by email here. |
Back to my homepage |
Last updated November 9, 2001.